by Richard
Haynes, Missouri USA
by Richard
Haynes, Missouri USA
Since childhood I have been intrigued by the marvelous color variety of
flowering plants: why and how do so many colors occur? Why so few blue
flowers and so many yellow? How do the flowers produce their colors and
of what substances are
they made? I had many questions but very few answers. [At this point I
must say I'm not a trained botanist or even a decent gardener. (My wife
has that attribute, thankfully.) I'm simply a retired chemist with
botanical questions.]
After earning a living in a commercial chemical direction that had
nothing to do
with gardening, botany or any of their aspects, I retired.
However, I was still interested in flower colors so I began a few
studies of yellow wild flowers because of their ubiquity and
ease of obtaining them. The flower I have studied the most has been a
Missouri region variety (Rudbeckia missouriensis)
of the Black-eyed Susan, Rudbeckia hirta.
R. missouriensis is a smaller
variety of its larger North American cousin, R. hirta, and has blossoms perhaps
one-half the size of the better known plant. Both are members of the
worldwide Daisy (asteraceae) family with its 1,000 genera and 20,000
species. Some other family flowers and plants are the sunflower,
coneflower, dandelion, coreopsis, yarrow, goldenrod, thistle, tansy,
etc., etc. [A botanical description of Rudbeckia missouriensis is at the end of this
article.]
|
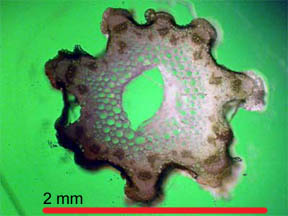
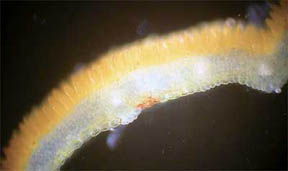
|
Fig. 1 R. missouriensis |
Fig.
2 R. missouriensis |
Extracting somewhat
pure, unchanged color pigments from wild flowers
is challenging, especially in a very small home laboratory not equipped
for organic synthesis. Once
extracted, examining spectra to attempt any identification is
also challenging, frustrating (especially without pigment comparisons)
and time consuming. So, I recently took time away from the project to
do other things. One of these was to use my microscope to have a go
at visually searching for the yellow pigments in the petals themselves.
(Actually, I thought this might be more entertaining rather than really
useful.)
I don't have a microtome but, by employing the ingenuous double razor
blade
method
reported by Walter Dioni (Micscape February 2004) I produced reasonably
useful slices. To maintain some cell osmotic pressure, the material was
cut under water (Dioni method) then mounted in weak saline solution
under cover glasses; I made no attempt at permanent mounts. All the
observations were essentially photographic and thus relatively brief.
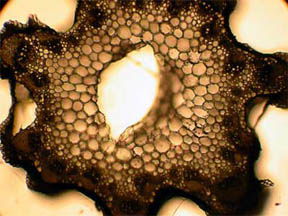
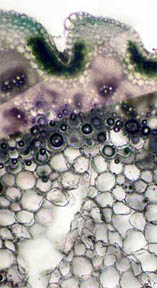
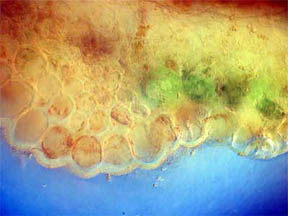
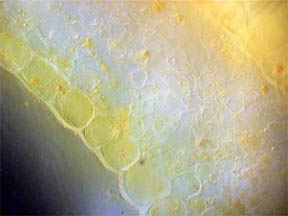
STEM
I looked first at transverse slices of
R. missouriensis stems that had
dried-on-the-stalk. (Here in Missouri we are in a summer-long
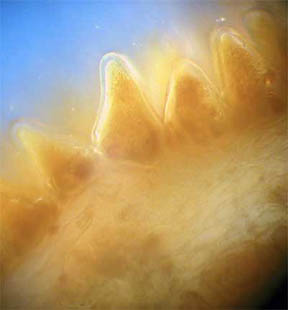
drought.) Figures 3 and 4, below, reveal a multi-sided, deep faceted
dry
stem with its central phloem core more than half gone and the small
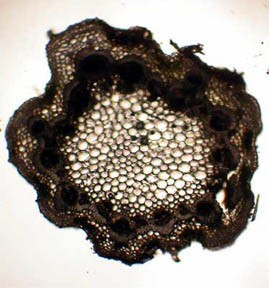
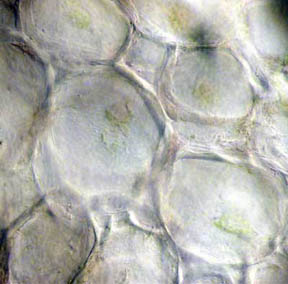
vascular bundles surrounding the stem are closed. Figures 5 and 6 of a
healthy stem, on the other hand, show that both the fluid bearing
central phloem cells and the vascular bundles are intact and
functioning. The greenish areas along the stem boundary, fig. 6,
contain many
chloroplasts in which chlorophyll is present. Two closer views of the
chlorophyll cell regions are seen in figures 7 and 8, both being
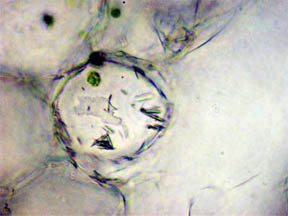
composites of more than one photo. Probably the needles seen
in the cells in figure 9 are
calcium oxalate (a metabolic
by-product) along with tiny
bundles
of chlorophyll-containing cells.
|
|
Fig. 3 Dry Stem, Rheinberg, 40X |
Fig. 4 Dry Stem, 100X |
|
|
Fig. 5 Fresh Stem, 40X |
Fig. 6 Rheinberg filter, 100X |
|
|
|
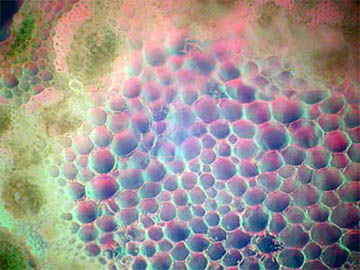
Fig. 7 Composite, 100X |
Fig. 8 Composite, 400X |
Fig. 9 Calcium Oxalate raphides, 400X |
A longitudinal slice (LS) of the dry stem, figures 10 and 11, also
reveals the loss of the central phloem cells. Bubbles in
Fig. 10 are from
water in the slide prep.
|
|
|
|
Fig. 10 LS, Dry Stem, 40X |
Fig. 11 LS, Edge Cells, 400X |
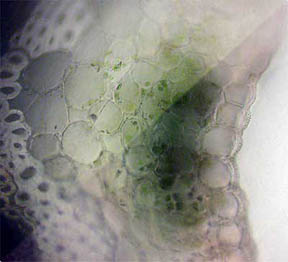
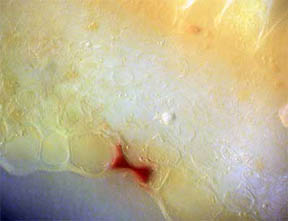
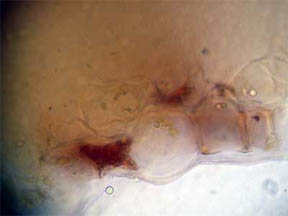
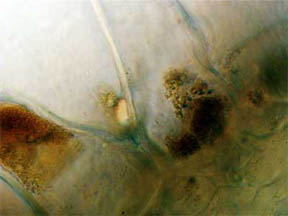
PERIANTH
A longitudinal slice through the perianth, the flower base
section composed of the sepals (calyx) and the petals (corolla), was
made to look for pigmented cells, especially where the petals attached.
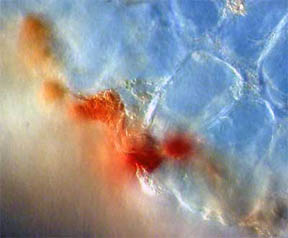
As might be expected, Fig. 12 and 13 at the bottom edge featured cells
filled with opaque red-orange pigment. Cells higher up the slice,
Fig. 14, contained areas of chlorophyll.
|
|
|
Fig. 12 Edge Cells of pigment, 100X |
Fig. 13 Closer view, 400X |
Fig. 14 Chlorophyll 400X |
PETAL SURFACE
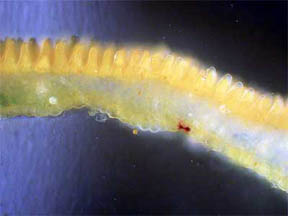
The surface of a few cut petals was examined in oblique
light; orderly rows of nipple-like (papillae) elongated surface cells
filled with
yellow pigment were seen. Figures 15 and 16 show the rows; figures 17
and 18 capture the papillae up close, 17 being very nearly a side view
while 18 is almost straight on. Papillae, which are small thickenings
of the cuticle and may be hollow or solid, often give a velvet
appearance to a petal. (The cuticle is a non-cellular outer protective
layer.)
|
|
Fig. 15 Surface papillae rows 40X |
Fig. 16 Closer example 100X |
|
|
Fig. 17 Side view of papillae 400X |
Fig. 18 Papillae almost straight on 400X |
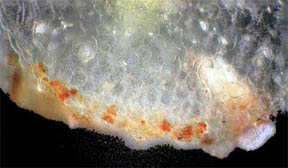
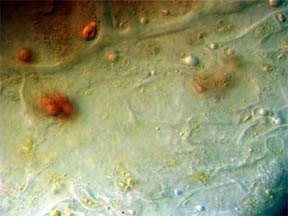
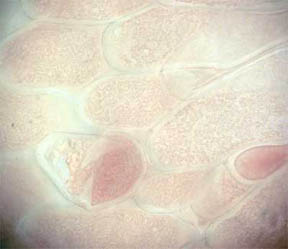
PETALS
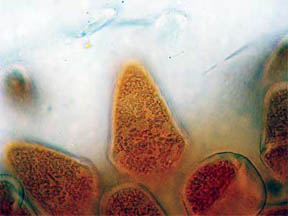
Finally, transverse sections of the petals themselves were put
under the 'scope. Flower petals are basically leaves that have
become transformed into the colorful but relatively fragile "glory of
the plant". Many petals of flowers show their leaf likeness by
possessing a few or many vein-like creases running through their
structures. Figure 19, below, is a TS of the central portion of a R. missouriensis petal. A crease or
dip in the section is evident. Figures 20 and 21 at 100X reveal small
pockets of red pigment along their bottom edges. Figure 22 at 400X
focuses on the bottom portion of the petal crease and shows both
chlorophyll cellular pockets as well as a rich scattering of red
pigment cells. Figures 23 and 24 continue the focus on the bottom edge
and the interior cellular structures with pigment packets very much in
evidence.
|
|
|
Fig. 19 Crease in petal 40X |
Fig. 20 Small red pigmented area 100X |
Fig. 21 Another pigment area 100X |
|
|
|
Fig. 22 Crease area with color cells near the bottom 400X |
Fig. 23 Red pigment bridging two edge cells (see fig. 20) 400X |
Fig. 24 Small Packets of pigment in cells 1,000X (oil) |
These areas of intense red-orange pigments most likely are carotenoid/xanthophyll compounds but may also be flavonoids. R. missouriensis contains both chemical groups though the carotenoids/xanthophylls seem to be present in larger amounts (my initial work appears to indicate this). In Nature, there are a number of yellow flowers that appear to be colored by some combination of carotenoid/flavinoid compounds. The carotenoids are oil soluble and are found in plastids within chromoplasts of the petals. (Interestingly, carotenoids often crystallize within chromoplasts.) Flavinoids are water soluble and are located in vacuole cavities. Anthrocyanins of the flavanoid family are present in many flowers and are the major contributors to red, purple and blue flowers. They too have a role in producing yellow in flowers but that role is as yet somewhat murky. And, the physical nature of the petal surface may likely have an effect on its color.
|
|
|
Fig. 25 Yellow pigment edge cells 400X |
Fig. 26 Pigments in bottom edge cells 1,000X (Oil) |
Fig. 27 Pigment in cells near the top surface in an apparent liquid state 1,000X (Oil) |
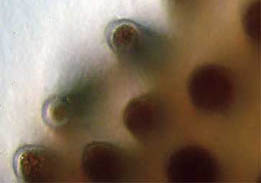
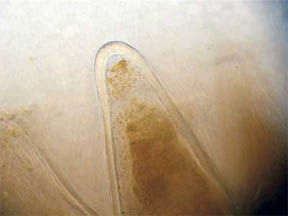
Observations of the papillae (below) were interesting. In one,
figure 28, most of the liquid formerly in the tips of the elongated
cells has disappeared leaving what appears to be a crystalline pigment
mass. The transparent nature of the cuticle tissue of the papillae is
quite distinct as well. Conversely, in figure 29 the entire papillae
are filled and a clear liquid in motion was seen churning around
the tip ends. And, while there may be some crystalline pigments,
most of the dark yellow mass appeared to be in solution. Figure
30 is a photo composite of the transverse slice, top to bottom. Because
of compositing, the photo is somewhat darker than the individual
pictures. However the petal structure difference between top and bottom
is clear. (The papillae themselves are quite small: their average
length is 0.077 mm and diameter is 0.027 mm.)
|
|
|
Fig. 28 Solid pigment 400X |
Fig. 29 Pigment in solution 400X |
Fig. 30 T/S composite 400X |
The papillae were looked at under highest magnification, figures 31 to 33, and more evidence of pigment clumping and/or crystallization was found in figure 32. In figure 31 the channel through which fluid flow was taking place is seen on the left side of the papilla and small amounts of pigment can be detected within the flow. Also the cuticle tip of the papilla is somewhat thicker than its walls and this was characteristic of most of the papillae observed. Almost all of the right papilla in figure 33 is empty of pigment though a mass of red-yellow pigment (?) sits turgidly at the bottom, seeming to mingle with chlorophyll containing cells.
|
|
|
Fig. 31 Pigment movement 1,000X (Oil) |
Fig. 32 Pigment clumping 1,000X (Oil) |
Fig. 33 Empty papilla 1,000X (Oil) |
At this point, I'm still studying my data and pictures to
determine if I've really learned anything about the yellow pigment
source(s) of Missouri's Black-eyed Susan, Rudbeckia missouriensis. Maybe yes, maybe no.
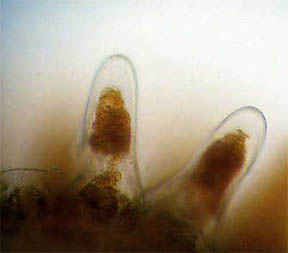
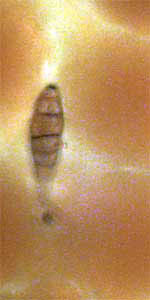
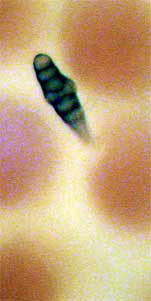
Oh, I almost forgot: I have a little puzzle for Micscape readers. In
looking over the petal surfaces at 400X, I discovered two tiny
critters, or I think they're critters, nestling between the papillae of
the upper surface. See figures 34 and 35. Both measure
approximately 0.01 mm in width and 0.04 mm in length. Any idea what
these unknown (to me, anyway) critters, bugs, cocoons, etc., might be??? Editor's
note added October 2006: Readers kindly emailed Richard to remark
on the nature of these organisms; they are 'Alternaria, a common fungal spore
that affects plants' and are described in the October
2006 issue.
|
|
Fig. 34 ??? 400X |
Fig. 35 ??? 400X |
I am interested to hear from Micscape readers and all comments are
welcome.
Missouri
Black-eyed Susan
- Rudbeckia missouriensis
Asteraceae
Daisy Family
Description: |
to 55 cm tall, branched, very hairy |
Flowerheads: |
terminal on a stalk, rays yellow
to yellow orange; 10 - 18 rays, often |
Leaves: |
linear with only the lowest ones lance-shaped |
REFERENCES
1. Raven, Peter H., Evert, Ray
F., Eichhorn, Susan E., Biology of Plants, 6th Ed.,
W.H. Freeman & Co., New York, 1999
2. Denison, Edgar, Missouri Wildflowers, 5th
Ed., MO Dept. of Conservation, Jefferson City, MO, 2001
3. Capon, Brian,
Botany for Gardeners,
Timber Press, Portland, OR, 1990
4. Goodwin,
T.W., Editor, Chemistry
& Biochemistry of Plant Pigments, Academic Press, London,
1965
A
note about the microscope and photos:
I
use a Nikon Eclipse 200 trinocular microscope equipped with an Qioptiq
digital coupler and a Nikon CP 4500 camera.
All photos were processed in Adobe Photoshop® 7.0 and some are of
differing size because I was looking for visual data and not
necessarily the best picture.
Published in the September 2006 edition of Micscape.
Please report any Web problems or offer general comments to the Micscape Editor .
Micscape is the on-line monthly magazine of the Microscopy UK web site at Microscopy-UK
© Onview.net Ltd, Microscopy-UK, and all contributors 1995
onwards. All rights reserved.
Main site is
at www.microscopy-uk.org.uk
with full mirror
at www.microscopy-uk.net
.